If you are in any way involved in biological recording as a volunteer, you’ll know the process – you see or collect a specimen and if you can identify it (or find someone you know who can), you write up the record in whichever format is appropriate, then send it off to the scheme organiser or records centre. In the UK, this is of huge importance – estimates vary, but the majority of biological records come from volunteers, from relative beginners to highly-experienced amateurs and professionals doing a bit extra in their spare time. If I remember rightly, around 90% of botanical records for the British ‘plant atlas’ were from volunteers with around 70% of records being collected voluntary for all species groups combined. As atlas publications need records, and conservation objectives are often based on biological data, it is clear how vital volunteer recorders are in Britain – some other countries such as France and Germany differ, with recording being undertaken mainly by paid workers, but focusing more on key policy-related species.
However, what happens when you get a specimen, lets say an invertebrate, that you can’t identify? You might know what family it’s in, or even genus, but the species eludes you. You look up the relevant recording scheme and get in touch with the scheme organiser – they may be able to offer advice, and might even be able to identify the beastie from a photo. However, if not (let’s say it needs dissection and more specialist references than you have access to), you might package it up and post it off to them. Well, in my ‘spare time’ I am one such such organiser, so I thought I’d share my view of this end of the process.
Not having had any recent enquiries, when a mystery Jiffy-bag landed on my doormat the other day, I was intrigued to find within a small plastic pot containing a small carded beetle and collection labels, a letter explaining its provenance and attempts to identify it and an address label and stamps to facilitate its return. Apart from not asking first (tut tut, a quick email to check availability is appreciated), this is exactly what I’m after – clear background info, and return postage for specimens.
A couple of days later I had time to look at it – described as having been swept from dune grassland in Cumbria during September, and identified by the sender as probably the genus
Longitarsus (a flea beetle within the family Chrysomelidae). My first thought (apart from ‘yes it is
Longitarsus’) was how neatly glue-mounted it was – at 2mm or so long, pinning would have been a challenge...
 |
| L. suturellus dorsal view - still mounted on its card. |
The sender had got stuck at species level, getting contradictory results from use of
Joy’s Practical Handbook (one of the beetle books, though many sections are now out of date) – particularly due to results indicating species not found in Cumbria. Now, I have written a key to the Chrysomelidae (though it’s not yet complete and is about to undergo its first major revision having been tested this summer). Working through this, I kept coming to the same result,
L. lycopi. Hmmm... that’s not right – not only is it hardly ever found in Cumbria (that doesn’t mean it’s wrong, but warrants closer checking), but despite the key, it didn’t look quite right (shape, colour etc). I had a feeling that the problem was with my key rather than the specimen (especially as I knew which species it looked like, but this was a good excuse to test the key further). So, going back to some other references, I realised which key couplet was the problem – the couplet was correct in itself, but needed further clarification regarding the sometimes fiddly process of looking at the lines and grooves on a tiny beetle’s ‘face’.
 |
| L. suturellus - the head in close-up. |
A tweak to the wording and the specimen consistently keyed out elsewhere – to
L. suturellus. This was much more promising (known from Cumbria), but as the genus is notoriously difficult to key to species using external features alone, the next step was dissection. It was a male, and teasing off the end of the abdomen using pins, the aedeagus could be extracted – it is distinctive in this species (somewhat like an ‘hourglass’ constriction with sturdy edges) and so the identification as
L. suturellus is confirmed and can be added to the scheme database. The specimen didn’t survive the dissection, but I did mount the aedeagus on a slide and sent that back to the collector who I hope will find it useful for reference.
 |
| Aedeagus of L. suturellus |
All fairly straightforward, and maybe an hour or so of effort, but it does highlight some important aspects of recording schemes such as this. Mostly, these are due to the fact that they are run by volunteers. Some organisers may work in closely related areas (e.g. as museum entomologists), but others do not. This isn’t necessarily a problem, but does mean that some aspects may be more difficult. For example, a wide range of sometimes obscure, sometimes expensive books and journal papers are likely to be required. Without ready access to an academic library or endless cash, this means reliance on interlibrary loans and the collections of record centres and museums. Now, I am lucky in that I am quite near Winchester where Hampshire’s excellent biological collection is housed. By appointment, I can use this collection (e.g. to check against known specimens), but (and this can be a large ‘but’), this has to be during ‘office hours’ i.e. when I tend to be at work... Fortunately my working life is quite flexible, but museum visits are still very limited, as is space and equipment for maintaining my own collection. Similarly, I am an inveterate bookworm, and have put a lot of time (and cash) into acquiring specialist papers and books, but still have a wish list of texts I haven’t been able to afford or obtain via inter-library loan (yet). Maybe I need to ‘monetise’ my blog!
Of course, recording schemes develop as does my involvement – I will continue to accumulate key texts, and the chrysomelid scheme should have a new website from spring 2011, hopefully with online recording. I also plan to develop a network of identifiers – at present, the scheme is still small as it was dormant when I took it over, but it will grow and with it the workload and the number of mystery Jiffy-bags. Apart from the nearly-grumble about museum access and book-money, I find it most satisfying – my entomological skills grow, and I know that the scheme will provide invaluable data for conservation and research, as well as (I hope) generating wider interest in the group as well as entomology and biological recording more broadly. For more, have a look at
my static page on the shiny, shiny Chrysomelidae, and I hope the boxes keep coming...
 |
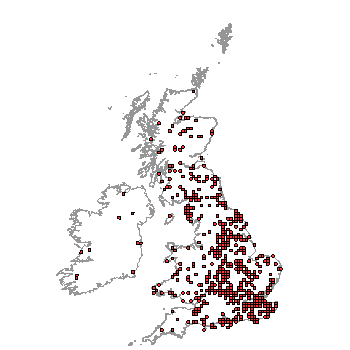
| NBN distribution map of L. suturellus - widespread! |