Lately I have written copiously about small invertebrates, particularly
those found recently in our firewood store. So, having written five parts of the woodpile series (so far), I felt it was time for a brief departure - in terms of both time and scale as I have decided to look at some aspects of the Cave Bear
Ursus spelaeus.
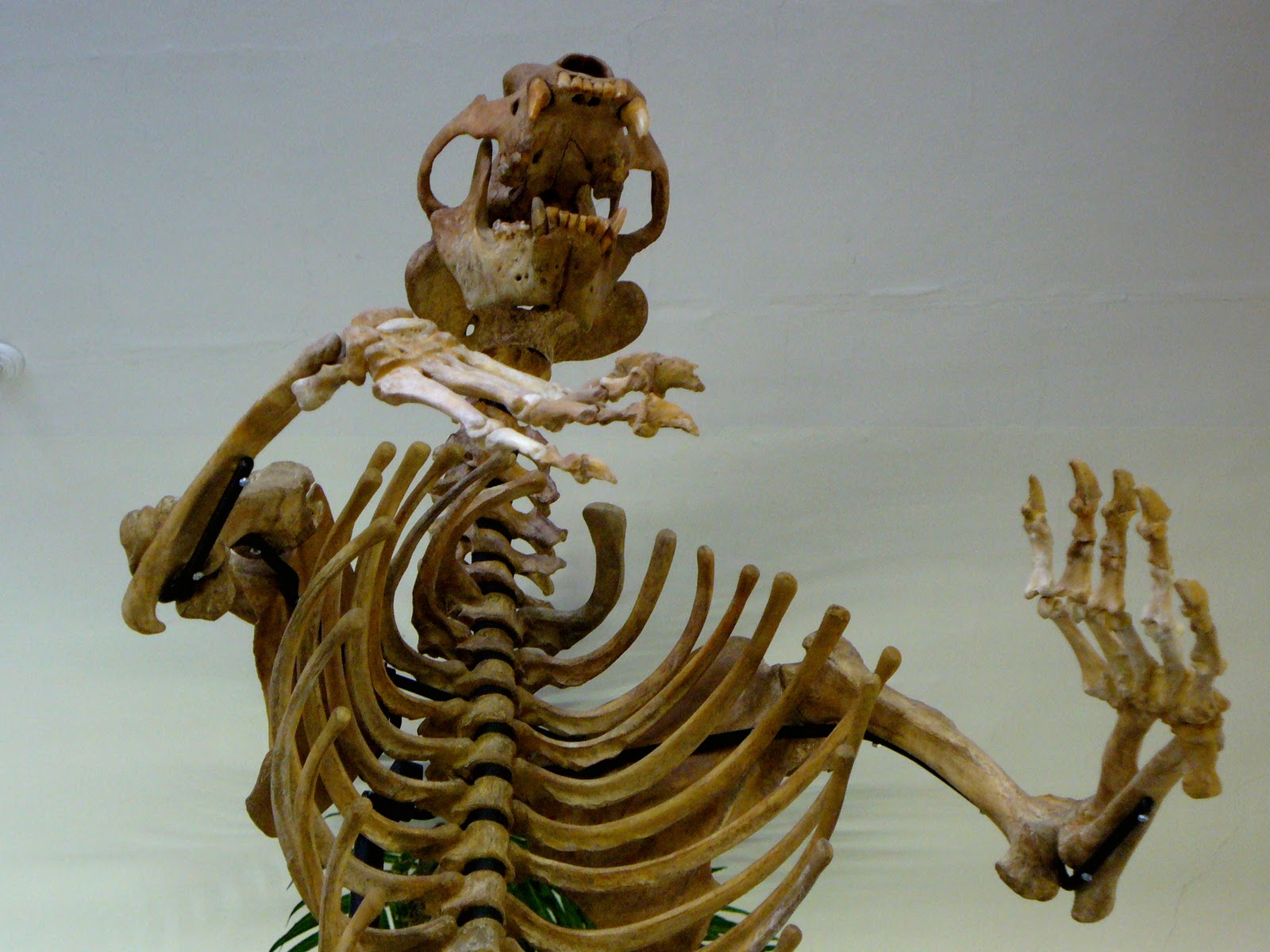
 |
| A Cave Bear skeleton in the typical (of museum displays) rearing posture. |
Cave Bears lived in Pleistocene Europe (the Pleistocene epoch lasted from around 2.5 million to 11,700 years ago and covers the most recent series of repeated glaciations) and current evidence suggests they became extinct around 27,800 years ago. This means that they would have been encountered by humans and indeed they are depicted in cave art (albeit rarely e.g. at
Les Combarelles cave in France). There is also possible evidence of Cave Bear worship by Neanderthals, such as at Drachenloch in Switzerland and Regourdou in France where the skulls of bears had clearly been arranged in and on man-made stone structures such as wall niches and a slab-covered pit. However, prehistoric anthropology, fascinating though it is, really isn't my area, so I'll stick with the more biological/ecological aspects. However, for an interesting overview of some aspects of human-Cave Bear interactions (focusing more on the earlier form of Cave Bear
U. deningeri which disappeared around 100,000 years ago and may be an earlier species, a transitional subspecies or simply a pre-interglacial form of U. spelaeus), have a look at
Stiner (1999). Taxonomic uncertainties aside, my interest was sparked when I bought a Cave Bear cheek tooth found in a cave in Romania, and dental evidence is where I will start.
 |
| Cave Bear cheek tooth, length 45mm. |
This tooth is in pretty good condition (it's still shiny after about 40,000 years which shows just how tough tooth enamel is) and has an extensive grinding surface with a couple of large bluntly pointed cusps. Cave Bears lost their premolars as they evolved, a feature which has been used to suggest a highly herbivorous diet (e.g. Kurtén, 1976). The last premolar evolved as a molar (molarisation) which allowed tough plant material to be chewed more effectively (and hence more more food energy to be extracted) due to the increased number of cusps and cutting edges of the teeth, especially in the elongated last molar. Their teeth also show more wear than in most modern bears which again suggests a herbivorous diet with a large component of tough/fibrous materials, although detailed analysis indicates that tubers and other gritty foods were not a major part of their diet, unlike for modern Brown Bears
U. arctos (Pinto Llona
et al., 2005). However, varying threads of research in this area, including evidence for some cannibalistic scavenging (Pacher, 2000) has led to current scientific opinion tending towards Cave Bears being more herbivorous than modern bears of the genus
Ursus, but still omnivorous to some extent. Recent re-examination of skull and tooth morphology (Figueirido
et al., 2009) and analysis of the regional variation in bone isotope composition, especially nitrogen-15 (Richards
et al., 2008; Trinkaus & Richards, 2008) both support this idea of omnivory and some variation in diet.
 |
| The same cheek tooth showing the pattern of the crown. The large grinding surface covers the left side of the tooth and the lower right side, with the pointed cusps to the upper right. The orange deposits in the grooves of the enamel are the remains of soil, although the right-hand end shows an area of worn (yellowish and not shiny) enamel at the base of the large cusp. |
With advances in molecular biological techniques, the possibility of investigating cave bear genetics arose and in 2005, nuclear DNA extracted from a Cave Bear tooth around 42-44,000 years old was sequenced. This indicated that the Cave Bear was more closely related to the Brown Bear and Polar Bear
U. maritimus than to the North American Black Bear
U. americanus (Noonan
et al., 2005) and supported earlier similar findings using mitochondrial DNA (Loreille
et al., 2001). Interestingly, investigation of the fine structure of Cave Bear tooth enamel (the 'rods' or 'prisms' that form the basic units of enamel) shows that it retained carnivore-like characteristics despite the clear adaptation to a largely herbivorous diet. Thus, changes in broad dental anatomy driven by dietary specialisation can occur without the equivalent changes in enamel structure (von Koenigswald, 1992), meaning that Cave Bears had herbivore-shaped cheek teeth with carnivore-like enamel.
So, we have an extinct species of bear clearly adapted to a specialised herbivorous diet with some elements of omnivory and variation. As well as the genetic evidence mentioned above, its skeleton is similar to that of the modern Brown Bear, with the two species appearing to have diverged around 1.2 to 1.4 million years ago (Loreille
et al., 2001) i.e. prior to the splitting of Brown and Polar Bears which may have occurred around 850,000 years ago, although this estimate is somewhat uncertain (Swenson, 2007). Males averaged 400–500 kg with females around half this weight at 225–250 kg (Christiansen, 1999), similar to the range for the largest modern bears, noting that they were larger during glaciations and smaller during interglacial periods (MacDonald, 1993), probably as an adaptation to adjust heat loss rate as larger animals have smaller surface area:volume ratios. The reason for its extinction is uncertain. It is unlikely to simply be due to its specialised diet and restricted geographical range ecologically 'marooning' it during post-glocial warming - after all, it had survived several similar changes in condition previously and there is possible genetic evidence of a decline starting some 25,000 years prior to its extinction (Stiller
et al., 2010). Also, as noted above there is strong evidence for the species' ability to vary its diet. Instead, it is likely that there was a complex interplay of factors, possibly involving competition with humans for cave habitat, maybe specifically for hibernation sites as Cave Bears did not appear to use alternatives such as forest thickets and failure to find a hibernation site would lead to death. Despite numerous media reports taking the 2010 paper by Stiller
et al. to be definitive evidence of competition with humans rather than changing climatic conditions to be the cause of Cave Bear extinction, there is still genuine scientific disagreement and research is ongoing. Further genetic work (Bon
et al., 2011) does however show reduced genetic diversity from specimens in France originating from the period directly prior to extinction (genetic diversity is greater for specimens prior to this), again indicating a species under stress during human colonisation of the area - and the possibility of competition for hibernation caves.
References
Bon, C., Berthonaud, V., Fosse, P., Gély, B., Maksud, F., Vitalis, R., Philippe, M., van der Plicht, J. & Elalouf, J.-M. (2011). Low regional diversity of late cave bears mitochondrial DNA at the time of Chauvet Aurignacian paintings.
Journal of Archaeological Science 38 (8): 1886-1895.
Christiansen, P. (1999). What size were
Arctodus simus and
Ursus spelaeus (Carnivora: Ursidae)?
Annales Zoologici Fennici 36: 93–102.
Figueirido, B., Palmqvist, P. & Pérez-Claros, J.A. (2009).
Ecomorphological correlates of craniodental variation in bears and paleobiological implications for extinct taxa: an approach based on geometric morphometrics. Journal of Zoology 277 (1): 70–80.
Kurtén, B. (1976).
The Cave Bear Story. Life and Death of a Vanished Animal. Columbia University Press, New York.
Loreille, O., Orlando, L., Patou-Mathis, M., Philippe, M., Taberlet, P. & Hänni, C. (2001). Ancient DNA analysis reveals divergence of the cave bear,
Ursus spelaeus, and brown bear,
Ursus arctos, lineages.
Current Biology 11 (3): 200
–203.
MacDonald, D. (1993).
The Velvet Claw: A Natural History of the Carnivores. BBC, London.
Noonan, J.P., Hofreiter, M., Smith, D., Priest, J.R., Rohland, N., Rabeder, G., Krause, J., Detter, J.C., Pääbo, S. & Rubin, E.M. (2005). Genomic Sequencing of Pleistocene Cave Bears. Science
309 (5734): 597
–599.
Pacher, M. (2000). Taphonomische Untersuchungen der Höhlenbären-Fundstellen in der Schwabenreith-Höhle bei Lunz am See (Niederösterreich). Beiträge zur Paläontologie 25: 11–85.
Pinto Llona, A.C., Andrews, P. & Etxeberrıa, P. (2005).
Taphonomy and Palaeoecology of Cave Bears from the Quaternary of Cantabrian Spain. Fondacion de Asturias/Du Pont Iberica/The Natural History Museum, Grafinsa, Oviedo.
Richards, M.P, Pacher, M., Stiller, M., Quilès, J., Hofreiter, M., Constantin, S., Zilhão, J. & Trinkaus, E. (2008). Isotopic evidence for omnivory among European cave bears: Late Pleistocene
Ursus spelaeus from the Peştera cu Oase, Romania.
Proceedings of the National Academy of Sciences of the United States of America 105 (2): 600–604.
Stiller, M., Baryshnikov, G., Bocherens, H., Grandal d'Anglade, A., Hilpert, B., Munzel, S.C., Pinhasi, R., Rabeder, G., Rosendahl, W., Trinkaus, E., Hofreiter, M. & Knapp, M. (2010). Withering Away
– 25,000 Years of Genetic Decline Preceded Cave Bear Extinction.
Molecular Biology and Evolution 27 (5): 975
–978.
Stiner, M.C. (1999). Cave bear ecology and interactions with Pleistocene humans.
Ursus 11: 41
–58.
Swenson, J.E. (2007).
Økologi hos en voksende bjørnebestand – Forvaltning når bjørnen har kommet tilbake. Det Skandinaviske Bjørneprosjektet [in Swedish] [accessed 15/11/2011].
Trinkaus, E. & Richards, M. P. (2008). Reply to Grandal and Fernández: Hibernation can also cause high δ15N values in cave bears. Proceedings of the National Academy of Sciences of the United States of America 105 (11): E15.
von Koenigswald, W. (1992). Tooth enamel of the cave bear (
Ursus spelaeus) and the relationship between diet and enamel structures.
Annales Zoologici Fennici 28: 217–227.